 2024,Vol. 33
2024,Vol. 33文章信息
- 赵常伟, 李传仁, 杨玉婷, 安维彬, 王在凌. 2024.
- ZHAO Changwei, LI Chuanren, YANG Yuting, AN Weibin, WANG Zailing. 2024.
- 不同寄主对南亚果实蝇生长发育和繁殖的影响
- Effect of different host plants on the growth, development and fecundity of Zeugodacus tau (Walker)
- 生物安全学报, 33(3): 229-235
- Journal of Biosafety, 33(3): 229-235.
- http://dx.doi.org/10.3969/j.issn.2095-1787.2024.03.004
文章历史
-
收稿日期(Received): 2023-06-24
接受日期(Accepted): 2023-07-19




2. 湖北省农林病虫害预警与调控工程技术研究中心, 湖北 荆州 434025;
3. 随州市曾都区农业局, 湖北 随州 441300
2. Forewarning and Management of Agricultural and Forestry Pests, Hubei Engineering Technology Center & College of Agriculture, Jingzhou, Hubei 434025, China;
3. Agricultural Bureau of Zengdu District, Suizhou City, Suizhou, Hubei 441300, China
南亚果实蝇Zeugodacus tau (Walker)属双翅目Diptera实蝇科Tephritidate,是我国一种非常重要的检疫性实蝇害虫(王奕婷等,2022)。南亚果实蝇主要分布在中国、日本、越南、菲律宾等亚热带及温带国家和地区(李春苑等,2012)。中国最早于1849在福建沿海地区发现其踪迹(汪兴鉴,1996)。因南亚果实蝇具有极强迁飞能力,且属于“r-对策”昆虫,种群数量快速扩增,现已在中国海南、广东、贵州、广西、重庆、湖北、湖南等16个省市区发生危害(王奕婷等,2022)。
南亚果实蝇寄主范围广,可危害葫芦科、茄科、番木瓜科和西番莲科等16科80多种寄主植物,充足的食物资源为南亚果实蝇种群的快速定殖提供了生存和繁殖基础(张艳和陈俊谕,2018)。南亚果实蝇以幼虫在果实内部取食,造成果实腐烂,提前脱落,导致严重减产。调查发现,南亚果实蝇对南瓜Cucurbita moschata (Duchesne ex Lam.) Duchesne ex Poir.的感染率最高,可达73.1%(李小珍, 2007),每年仅对南瓜产业造成的潜在经济损失可达3千万~2亿元(方焱等,2015)。
多食性昆虫对寄主的寻找和取食过程主要受寄主植物营养成分、挥发性气味成分的影响,因而对寄主产生不同嗜好性(卢伟,2007;钦俊德, 1996)。不同寄主间、同一寄主植物不同年份和不同季节间以及不同立地环境之间的基础营养与次生代谢物质等组分上均存在明显差异,继而影响昆虫的发育进度、存活及繁殖能力(Heindl et al., 2023;Zhao et al., 2023)。因此,昆虫对不同寄主的适应性差异是影响昆虫种群数量的重要因素,直接影响植食性昆虫的生长发育、成虫寿命和繁殖能力(钦俊德, 1996)。
目前,南亚果实蝇在不同寄主植物上的适应性研究主要集中于葫芦科的冬瓜Benincasa hispida (Thunb.) Cogn.、南瓜等寄主植物,在茄科植物上的研究较少。湖北地区瓜果类、茄科类蔬菜产量分别占总蔬菜产量的11.08%、10.66 %,是湖北省非常重要的经济果蔬(胡世霞,2016))。因此,本试验选取葫芦科植物南瓜、黄瓜Cucumis sativus L.、冬瓜和茄科植物西红柿Solanum lycopersicum L.为试验材料,通过两性生命表技术评价南亚果实蝇在不同寄主植物上的存活率和繁殖力,明确南亚果实蝇在不同寄主上的繁殖优势,为江汉平原地区田间寄主植物种植结构的调整及绿色防治提供理论依据。
1 材料和方法 1.1 供试昆虫南亚果实蝇采集自湖北省荆州市长江大学西校区(112°23′74″N,30°35′75″E)南瓜田中。将有南亚果实蝇幼虫的南瓜放置于温度(25±1) ℃、相对湿度(75±5)%、光周期12 L: 12 D的恒温培养箱(GZX-400BS-Ⅲ,宁波赛福实验仪器有限公司)中。待蛹羽化为成虫并准确鉴定后,分别以南瓜、黄瓜、冬瓜和西红柿饲养3代以上。
1.2 供试寄主长江大学西校区附近菜市场采购新鲜且成熟的南瓜、黄瓜、冬瓜和西红柿。采购回来的南瓜、黄瓜、冬瓜和西红柿使用去离子水清洗3遍后,置于4 ℃冰箱保存,待后续试验使用。
1.3 南亚果实蝇室内种群试验将不同寄主饲养的第3代南亚果实蝇成虫分别放入4个饲养笼(40 cm×40 cm×40 cm)中,每个饲养笼中约20对南亚果实蝇成虫。将南瓜、黄瓜、冬瓜和西红柿切成长约10 cm,宽约5 cm的小块,每小块果肉切开4~5条深度为1~2 cm的裂缝后,放入装有相应寄主的饲养笼中,供南亚果实蝇成虫产卵。2 h后拿出带有南亚果实蝇卵粒的果肉,置于温度(25±1) ℃、相对湿度(75±5) %、光周期12 L∶12 D的恒温培养箱(GZX-400BS-Ⅲ)中。待卵孵化后用细毛笔随机挑选100头活跃初孵化幼虫至一次性培养皿(直径4 cm、高1 cm)中单头饲养,每天更换寄主食物,在体视镜下观察并记录龄期(幼虫龄期以蜕皮为标准)和存活情况。
幼虫发育至3龄老熟幼虫时(频繁上下跳动为老熟幼虫的标志),将老熟幼虫置于厚度2.5 cm、相对湿度(75±5) %的灭菌沙表面,供老熟幼虫化蛹。待老熟幼虫入土后第4天,取出所有的蛹,使用软毛刷清除周围沙粒后用卫生纸将蛹表面的水分擦干。在千分之一天平(OHAUS-AX224ZH/E)称重后放回塑料杯中,盖上0.5~1.0 cm厚的沙粒。
南亚果实蝇蛹羽化后,将同种寄主饲养的雌雄成虫进行配对,并置于放有10%蜂蜜水的饲养笼中,分别使用长约10 cm、宽约5 cm的南瓜块、冬瓜块、西红柿块和黄瓜块引诱南亚果实蝇产卵,每天记录产卵量、卵孵化率和成虫存活情况。
1.4 种群生命表的构建及种群动态预测参照Chi & Liu (1985)及Chi & Getz (1988)提出的年龄-阶段两性生命表研究方法,建立南亚果实蝇在4种寄主寄主植物上的种群生命表。sxj表示南亚果实蝇在年龄x、阶段j的存活率,lx为忽略龄期的情况下种群在年龄x的存活率(齐心等,2019;Chi et al., 2020)。两者之间的关系为:
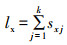
|
种群特征繁殖力mx是指所有存活的南亚果实蝇在年龄x、阶段j时的平均产卵量,年龄-阶段特征繁殖力(fx)是指在年龄x、阶段j时所有存活雌虫的平均产卵量。二者之间的计算公式为:
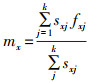
|
净增长率R0是南亚果实蝇种群个体从出生到死亡的存活率(lx)与种群特征繁殖力(mx)乘积之和:
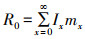
|
内禀增长率r的计算公式:

|
周限增长率λ的计算公式:
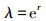
|
使用年龄-龄期两性生命表软件TWOSEX-MSChat (Chi,2020a)分析和计算南亚果实蝇在4种寄主植物上的年龄-龄期存活率(sxj)、年龄特征存活率(lx)、年龄特征繁殖力(fx)、种群特征繁殖力(mx)、种群特征净增值率(lxmx)、内禀增长率r、周限增长率λ、净增值率R0和平均世代周期T。TIME-MSChat软件(Chi,2020b)预测南亚果实蝇在4种寄主上发育140 d后的种群数量。南亚果实蝇在4种寄主植物上的发育历期、成虫寿命、繁殖力等生活史参数的标准误均选择Bootstrap重复100000次进行计算,通过Paired bootstrap test (5%)对南亚果实蝇在4种寄主上的生活史参数进行差异性检验。
2 结果与分析 2.1 寄主食物对南亚果实蝇发育历期、蛹重和存活率的影响南亚果实蝇在冬瓜上的1龄(2.41 d)、2龄(2.58 d)和3龄(3.22 d)幼虫的发育历期以及整个幼虫期(8.49 d)均显著长于在南瓜、黄瓜和西红柿上的发育历期。南亚果实蝇1、2、3龄幼虫的发育历期以及整个幼虫期在南瓜、黄瓜和西红柿上无显著差异。在4种寄主上的蛹期无显著差异。取食西红柿的蛹重(16.28 mg)显著高于取食其他3种寄主,取食冬瓜的蛹重(10.78 mg)显著低于取食其他3种寄主食物。南亚果实蝇在4种寄主植物上的成虫前期存活率存在显著差异,其中,在南瓜上成虫前期存活率最高(87.96%),显著高于取食其他3种寄主植物的成虫前期存活率(表 1)。
| 寄主Host | 1龄1st instar/d | 2龄2nd instar/d | 3龄3rd instar/d | 幼虫期Total larval stage/d | 蛹期Pupal stage/d | 蛹重Pupa weight/mg | 成虫前期存活率Pre-adult survival rate/% |
| 南瓜 | 2.10±0.04b | 2.08±0.10b | 2.71±0.07b | 6.74±0.10b | 8.31±0.06a | 14.59±0.54b | 87.96±3.23a |
| Cucurbita moschata | |||||||
| 黄瓜 | 2.12±0.06b | 2.14±0.04b | 2.84±0.11b | 6.98±0.15b | 8.39±0.03a | 13.75±0.47b | 80.00±4.00b |
| Cucumis sativus | |||||||
| 冬瓜 | 2.41±0.10a | 2.58±0.05a | 3.22±0.03a | 8.49±0.09a | 8.42±0.17a | 10.78±0.83c | 72.03±4.47c |
| Benincasa hispida | |||||||
| 西红柿 | 2.14±0.02b | 2.14±0.02b | 2.98±0.06b | 7.16±0.04b | 8.29±0.04a | 16.28±0.34a | 69.69±4.62d |
| Solanum lycopersicum | |||||||
| 表中数据为平均值±标准误,同一列数据后不同小写字母表示使用bootstrap (B=100000)程序检验在0.05水平差异显著。 Data are mean±SE. The different lowercase letters in the same column indicate significant differences at 0.05 level (paired bootstrap test, B=100000). | |||||||
幼虫取食冬瓜, 产卵前期(25.24 d)和总产卵前期(41.37 d)显著长于寄主食物为南瓜(20.71 d、36.77 d)和黄瓜(21.20 d、36.58 d)的产卵前期和总产卵前期。幼虫取食南瓜, 产卵期(50.25 d)、雌虫寿命(101.36 d)、雄虫寿命(141.89 d)显著长于取食其他3种寄主植物的产卵期、雌虫寿命和雄虫寿命。幼虫取食南瓜,其成虫的产卵量(361.26粒)显著高于取食其他3种寄主寄主的成虫产卵量。卵孵化率在南瓜(80.81%)和黄瓜(79.21%)之间无显著差异,但显著高于取食冬瓜(67.61%)和西红柿(72.58%)的卵孵化率(表 2)。
| 寄主Host | 产卵前期Adult preoviposition period/d | 总产卵前期Total preoviposition period/d | 产卵期Oviposition period/d | 雌成虫寿命Female longevity/d | 雄成虫寿命Male longevity/d | 产卵量/粒Fecundity | 卵孵化率Hatching rate/% |
| 南瓜 | 20.71±1.23b | 36.77±1.05b | 50.25±6.06a | 101.36±15.23a | 141.89±10.22a | 361.26±23.11a | 80.81±0.27a |
| Cucurbita moschata | |||||||
| 黄瓜 | 21.20±1.76b | 36.58±2.57b | 44.13±8.04b | 70.78±18.55b | 84.61±9.55b | 288.34±23.56b | 79.21±0.16a |
| Cucumis sativus | |||||||
| 冬瓜 | 25.24±1.59a | 41.37±2.17a | 32.76±7.38c | 43.08±8.22c | 57.53±8.42c | 84.80±11.01c | 67.61±0.18b |
| Benincasa hispida | |||||||
| 西红柿 | 23.75±1.01ab | 39.75±0.68ab | 34.64±6.42c | 47.92±12.66c | 39.31±7.17d | 76.55±9.64c | 72.58±0.19b |
| Solanum lycopersicum | |||||||
| 表中数据为平均值±标准误,同一列数据后不同小写字母表示使用bootstrap (B=100 000)程序检验在0.05水平差异显著。 Data are mean±SE. The different lowercase letters in the same column indicate significant differences at 0.05 level (paired bootstrap test, B=100000). | |||||||
南亚果实蝇在南瓜、黄瓜、冬瓜和番茄上的幼虫期年龄-龄期存活率分别是0.88%、0.86%、0.74%和0.81%,但南亚果实蝇在南瓜(0.85%)、黄瓜(0.78%)、冬瓜(0.72%)和番茄(0.69)上的成虫最高年龄-龄期存活率sxj均低于同种寄主食物下幼虫期的年龄-龄期存活率(图 1、2)。南亚果实蝇取食南瓜,其整个生命周期(卵期+幼虫期+蛹期+成虫期)最长的可达208 d,而南亚果实蝇取食冬瓜和西红柿,其成虫的最长生命周期仅有128和109 d(图 1、2)。取食南瓜的南亚果实蝇雌雄虫的存活率在30~130 d时缓慢下降,而在130 d后,南亚果实蝇雌雄虫的存活率急剧下降,且雌虫存活率的下降速率快于雄虫存活率的下降速率。取食黄瓜和西红柿的南亚果实蝇成虫存活率在30 d后快速下降,同时雌虫的死亡速度与雄虫的死亡速度相当。且取食西红柿的南亚果实蝇成虫存活率下降速率快于取食黄瓜的南亚果实蝇(图 1、2)。

|
图 1 南亚果实蝇卵、幼虫和蛹在4种寄主上的年龄-龄期存活曲线 Fig.1 Age-age survival curves (sxj) of egg, larvae and pupae of Z. tau on four hosts |

|
图 2 南亚果实蝇雌雄虫在4种寄主上的年龄-龄期存活曲线 Fig.2 Age-age survival curves (sxj) of emale and male of Z. tau on four hosts |
南亚果实蝇在4种寄主果实上的年龄-龄期特征存活率(lx)、年龄-龄期特征繁殖力(fx)和种群特征繁殖力(mx)如图 3所示。南亚果实蝇的繁殖力曲线由多个不连续的高低峰接连而成,且存在间断不产卵的情况。南亚果实蝇幼虫取食南瓜,其成虫的年龄-龄期特征繁殖力(14.03)在43 d时达到最高,高于取食黄瓜(10.26)、西红柿(10.09)和冬瓜(9.78)的的年龄-龄期特征繁殖力。南亚果实蝇在4种寄主上的年龄-龄期特征存活率lx随着龄期的增长逐渐下降。

|
图 3 南亚果实蝇在4种寄主上的年龄-特征存活率和种群繁殖力 Fig.3 Age-specific survival rate and fecundity of Z. tau on four hosts |
南亚果实蝇在4种寄主上的种群参数如表 3所示。南亚果实蝇在南瓜上的内禀增长率(0.10)和净增值率(90.63)显著高于在黄瓜(0.08、74.37)、冬瓜(0.07、35.62)和西红柿上(0.07、26.03)的内禀增长率和净增值率。南亚果实蝇在4种寄主上的周限增长率无显著差异。南亚果实蝇在黄瓜(54.50 d)和冬瓜(56.92 d)的平均世代周期显著长于在西红柿(48.41 d)上的平均世代周期。
| 寄主Host | 内禀增长率Intrinsic rate of increase | 净增值率Net reproductive rate | 周限增长率Finite rate of increase | 世代平均周期Mean generation time/d |
| 南瓜Cucurbita moschata | 0.10±0.01a | 90.63±8.25a | 1.10±0.02a | 49.89±2.24ab |
| 黄瓜Cucumis sativus | 0.08±0.01b | 74.37±4.47b | 1.08±0.01a | 54.50±4.51a |
| 冬瓜Benincasa hispida | 0.07±0.00b | 35.62±6.21c | 1.06±0.05a | 56.92±2.05a |
| 西红柿Solanum lycopersicum | 0.07±0.00b | 26.03±6.88c | 1.06±0.01a | 48.41±1.07b |
| 表中数据为平均值±标准误,同一列数据后不同小写字母表示使用bootstrap (B=100000)程序检验在0.05水平差异显著。 Data are mean±SE. The different lowercase letters in the same column indicate significant differences at 0.05 level (paired bootstrap test, B=100000). | ||||
南亚果实蝇在4种寄主植物上发育140 d的种群预测数量和种群预测动态如图 4所示。南亚果实蝇在南瓜上饲养的第二代新生卵的种群数量高于其他3种寄主饲养的新生卵数量。在140 d时南亚果实蝇在南瓜上的种群数量指数最高(5.72),其次为黄瓜(4.36),最低为冬瓜(3.56)和西红柿(3.42)。以南瓜和黄瓜为寄主饲养的南亚果实蝇在85 d后可以发育第3代成虫,而以冬瓜和西红柿饲养需95 d才能与羽化出第3代。在140 d时,以南瓜和黄瓜饲养的南亚果实蝇第4代成虫已经开始羽化,而以冬瓜和西红柿饲养的第3代成虫还未完全羽化。

|
图 4 南亚果实蝇在4种寄主上繁殖的种群动态预测 Fig.4 Prediction of population dynamics of Z. tau fed on four hosts |
与传统生命表相比,年龄-龄期两性生命能更清楚地反映种群在不同环境、不同寄主条件下的种群生长发育差异,是评价昆虫种群寄主适应性及对未来种群数量预测的重要方法。周波(2005)通过传统生命表记录南亚果实蝇在南瓜和黄瓜上的幼虫发育历期和存活率,与本试验结果接近,但在南瓜和黄瓜上的单雌产卵量仅为84.67和99.56粒,显著低于本试验结果。该差异可能是由于周波(2005)使用3%的蜂蜜水饲喂成虫,成虫期的营养不足,从而导致成虫产卵量显著下降。
南亚果实蝇在南瓜、黄瓜和西红柿上幼虫的发育历期无显著差异,但是以西红柿为寄主的蛹重(16.28 mg)显著高于取食冬瓜和南瓜的蛹重。李丽丽(2009)发现橘小实蝇Bactrocera dorsalis (Hendel)幼虫取食碳水化合物和蛋白质的比例和含量显著影响幼虫生长发育和蛹重的大小。由此推测,西红柿中碳水化合物和蛋白质含量可能更有利于南亚果实蝇幼虫的生长发育。
南亚果实蝇在武汉地区的发生高峰期为6—9月,此阶段为寄主果实逐渐成熟的时期且温度较高,利于其存活和繁殖(张振宇等,2018)。本试验中南亚果实蝇在4种寄主上的总产卵前期无显著差异,均在20 d后繁殖下一代。在无外界条件限制下,预测140 d后以南瓜为寄主的雌成虫数量可达50万头,以西红柿为寄主的雌成虫数量仅有2682头。因此,在田间应调整种植结构, 避免南亚果实蝇危害葫芦科的冬瓜、黄瓜后,转移危害茄科作物。
本试验首次使用两性生命表技术评价南亚果实蝇在4种寄主上的适合度,确定南瓜是南亚果实蝇最适宜的寄主,同时明确南亚果实蝇可以在冬瓜、黄瓜和西红柿上完成整个世代周期。本试验南亚果实蝇成虫各阶段发育历期、产卵高峰期等生物学参数可为江汉平原地区南亚果实蝇的防治提供生物学依据。
方焱, 李志红, 秦萌, 吴志刚, 赵守歧, 吴立峰, 赵中华, 陈克, 秦誉嘉, 王聪, 赵谈. 2015. 南亚果实蝇对我国南瓜产业的潜在经济损失评估. 植物检疫, 29(3): 28-33. |
胡世霞, 2016. 湖北省蔬菜产业竞争力研究. 硕士学位论文. 武汉: 华中农业大学.
|
李春苑, 吴佳教, 胡学难, 刘海军. 2012. 南亚果实蝇各虫态发育历期及有效积温研究. 生物安全学报, 21(2): 105-108. DOI:10.3969/j.issn.2095-1787.2012.02.005 |
李丽丽, 2009. 橘小实蝇大量饲养技术优化. 硕士学位论文. 福州: 福建农林大学.
|
李小珍, 2007. 南亚果实蝇种群特征及其对食料和热胁迫的生理调控机制. 硕士学位论文. 重庆: 西南大学.
|
卢伟, 2007. 植物营养对烟粉虱行为、发育和种群参数的影响. 硕士学位论文. 长沙: 湖南农业大学.
|
钦俊德. 1996. 昆虫与寄主植物的适应性及协调进化. 生物学通报, 31(1): 1-3. |
齐心, 傅建炜, 尤民生. 2019. 年龄-龄期两性生命表及其在种群生态学与害虫综合治理中的应用. 昆虫学报, 62(2): 255-262. |
王奕婷, 白强, 陈红松, 田震亚, 高旭渊, 周忠实. 2022. 瓜实蝇和南亚果实蝇在我国的分布差异. 环境昆虫学报, 44(5): 1170-1175. DOI:10.3969/j.issn.1674-0858.2022.05.11 |
汪兴鉴. 1996. 果实蝇属重要害虫的厘订名录和分亚属及种检索表(双翅目: 实蝇科: 寡鬃实蝇亚科). 植物检疫, 10(2): 95-103. |
张艳, 陈俊谕. 2018. 南亚果实蝇国内研究进展. 热带农业科学, 38(11): 70-77. |
张振宇, 李亮, 邓义, 焦丽, 凌霄霞, 张宏宇. 2018. 湖北武汉果实蝇害虫种群动态调查. 华中农业大学学报, 37(5): 52-58. |
周波, 2005. 南亚实蝇生物学特性及不同食料对其种群增长的影响. 硕士学位论文. 重庆: 西南农业大学.
|
CHI H, 2020a. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. . http://140.120.197.173/Ecology/.
|
CHI H, 2020b. TIMING-MSChart: a computer program for the timing of pest management. . http://140.120.197.173/Ecology/.
|
CHI H, LIU H. 1985. Two new methods for the study of insect population ecology. Academia Sinica, 24(2): 225-240. |
CHI H, YOU M, ATLHAN R, SMITH C, KAVOUSI A, ÖZGÖKCE M, GUNCAN A, TUAN S J, FU J W, XU Y Y. 2020. Age-stage, two-sex life table: an introduction to theory, data analysis, and application. Entomologia Generalis, 40(2): 102-123. |
CHI H, GETZ W M. 1988. Mass rearing and harvesting based on an age-stage, two-sex life table: a potato tuberworm (Lepidoptera: Gelechiidae) case study. Environmental Entomology, 17(1): 18-25. DOI:10.1093/ee/17.1.18 |
HEINDL M R, BÖTTCHER-FRIEBERTSHÄUSER E. 2023. The role of influenza-A virus and coronavirus viral glycoprotein cleavage in host adaptation. Current Opinion in Virology, 58: 101303. |
ZHAO J S, YAO Y F, DONG M M, XIAO H T, XIONG Y, YANG S Z, XU H L. 2023. Diet and high altitude strongly drive convergent adaptation of gut microbiota in wild macaques, humans, and dogs to high altitude environments. Frontiers in Microbiology, 14: 1067240. |